研究紹介
水の地球化学からハイドレートの挙動を探る
堆積物粒子間の水(間隙水)に溶けているイオンは、ガスハイドレートの生成と密接にかかわっている。メタンハイドレートの生成にはメタンが最重要の要素であり、堆積物粒子間の間隙水が溶存メタンに飽和する必要がある。酸化的な海底面付近でメタン飽和状態に達するには、酸化によるメタンの消費を避けるために、間隙水が還元的な環境に変化する必要がある。ガスハイドレートと間隙水イオンの関わりの一つが、還元的環境の発達による間隙水の組成変化である。2つ目は、すでに還元的環境になっている海底面深部、あるいは還元的環境形成後のメタンハイドレートの発達による組成変化である。
還元的環境の発達に伴う間隙水の化学組成変化
間隙水の化学組成は、酸化的な海水に接している海底面から、少しずつ変化している。これは、微生物による酸化・還元反応(酸化剤を使った有機物の分解)が、徐々に変化するからである。そして、間隙水にメタンが存在できるようになるまでには、間隙水中に硫酸イオン(SO42-)が枯渇した状態にまで変化する必要がある。SO42-を使った嫌気的メタン酸化(AOM)によって生じるエネルギーは最も小さく、他の酸化剤(酸素; O2、マンガン; Mn、鉄; Fe、硝酸; NO3など)が順に消費された後で、ようやく始まる。
間隙水がSO42-を含む所を硫酸帯(Sulfate zone)、それより深くメタンが存在する部分をメタン帯(Methanogenesis zone)と呼んで区別する。その境界は一部重なっており、SO42-とメタンが共存している部分を硫酸―メタン トランジッション(SMT)と呼ぶ(トランジッションの厚さが薄く、無視できるとした場合には、硫酸―メタン インターフェイス(SMI)と呼ぶこともある)。SMTの深さは、地下深部からのメタンの供給量(フラックス)を反映していると考えられている (e.g., Borowski et al., 1996)。これは、①海水中のSO42-濃度に変化が無いことから、SO42-は制限要素にならないこと、②メタンとSO42-の両方が必要なAOMがSMTでしか起こらない反応であり、SMTの形成が、いずれの材料とも供給過多になっていないためである。メタン帯の発達(海底面へ向かって広がる現象)は、メタンの供給量が増え、SMT深度が浅くなり、硫酸帯が減少につながる。Methanogenesis zoneが、海底面付近のFeやMnの酸化・還元帯に接近・重複すれば、酸化されたFeやMnを使ったAOMがFeやMnを溶かし、間隙水に影響を与えられる。酸化Feは、希土類元素(REE)のキャリヤーである (Haley et al., 2004)ため、メタンフラックスは、REEの挙動にも影響を与えられる。
メタンハイドレートの生成に伴う間隙水の化学組成変化
メタンハイドレートは、電荷をもった物質を嫌う。そのため、ガス分子は結晶中に取り込まれるが、電荷を帯びたイオンは排除される。この現象は、ion exclusionと呼ばれている(Ussler and Paull, 2001)。活発なハイドレートの生成が起こっている場所では、間隙水中の塩素イオン(Cl-)が増加する現象がみられる。Cl-が使われるのは、このイオンが、最も化学活性が低いからである(その他のイオンは、堆積物中の鉱物との反応があるため、濃度異常があっても、原因の説明が単純ではない)。ハイドレートの生成が起こっていた場所では、イオンの拡散により、この異常は小さくなり(海水の値に近づき)、やがては痕跡が残らなくなる。
メタンハイドレートの生成は、イオンの濃度以外にも異常を引き起こす。メタンハイドレートの格子に使われる水分子は、安定化のために、同位体組成の高い分子が選択的に利用される(水分子に関する同位体分別)。結果として、メタンハイドレートの周囲には、水素・酸素同位体比の軽い水分子が取り残される。
メタンハイドレートが分解した場合には、逆の現象(同位体比の高い水の放出による、イオン濃度の低下と、同位体組成の増加)が起こるが、その程度は、生成に起こった異常が、ハイドレートの分解開始までにどれだけ消失していたかによる。
メタンハイドレートは、生成・分解のいずれの場合も間隙水化学組成に影響を与えるため、メタンハイドレートを含んだ泥試料(あるいは含んでいたと考えられる試料)を回収した場合には、間隙水の化学組成の解釈が単純にはいかない。さまざまな情報を組み合わせて解釈する必要がある。
底生生物群集による環境復元の方法
1)ハイドレート産出域における現生底生有孔虫群集の解析
1-1.はじめに
これまで日本周辺海域の日本海や北西太平洋では,いくつかの海域でメタンハイドレートの存在が報告されています.我々の研究グループでは,日本海の上越沖海域を中心に2004年から,研究調査船による海底調査を実施し,ガスチムニー構造・メタンプルーム・海底地形の異常に基づいた表層型ガスハイドレートの回収・観察・分析を行っています (Matsumoto et al., 2005).その一方で,メタンハイドレートの開発のためには,掘削に伴う環境影響評価(メタンガスや採掘時のカッティングスが及ぼす生物環境変化)について考慮する必要があり,まず本年度では,現在の海底環境や生物環境について環境ベースライン調査および解析を実施しています.
1-2.有孔虫とは・・
私が研究テーマとしている有孔虫とは,海洋中に豊富に幅広く生息している単細胞生物で,かたい殻が大昔の化石としても大量に保存されているため,過去の地層年代や海洋環境を知るうえで有用です.また,数多くの殻形態の異なる種が知られており,それぞれが様々な海洋環境の違いに適応して棲み分けを行っています.北太平洋域では,体内にメタン酸化細菌や硫黄酸化細菌を細胞の中に共生させている特別な種の存在が確認されています.
1-3.ハイドレート産出域における底生環境の復元
日本海のメタンハイドレートが産出する場所では,所々で海底下よりメタンガスが湧出しており,ズワイガニやシロウリガイなどの特殊な大型動物が生息しています.しかしながら,微小単細胞生物の有孔虫についてはまだはっきりとわかっていません.私たちは,ROV「ハイパードルフィン」支援母船「なつしま」調査潜航調査航海において,日本海上越沖などのメタン湧出地域よりMBARI式プッシュコアラーを用い長さ数十cmの海底柱状堆積物試料を採取し,染色された有孔虫の生体個体を実体顕微鏡下で観察しています.これらの有孔虫の生息分布結果(有孔虫フラックスおよび底生有孔虫群集組成)は,現在のメタンハイドレート海域における生物環境のベースライン調査に役立っています.今後は,メタンハイドレートの掘削に伴う生物環境変化を明らかにするため,掘削前後における海底堆積物中の有孔虫組成を半年~1年間隔で追跡していくつもりです.
2)メタンハイドレート域で採取された海底コアにおける微化石調査
2-1.はじめに
ガスハイドレート研究所では,メタンハイドレート分布域で複数の海底コアを採取し,ハイドレート形成域における地質学的解析を実施しています.私たちの層序学グループでは,さまざまな古環境指標から導かれる堆積層序や微化石層序の結果に基づき,過去のハイドレート生成年代や生成時の海底環境などを予測しています.特に,上越沖における堆積層序や有孔虫化石や珪藻化石の微化石層序がすでに明らかにされているため,それらの層序帯と比較することですばやく地質年代や古環境を推定できます.また,有孔虫化石を拾いだし放射性炭素(14C)年代と安定炭素酸素同位体(d18O&d13C)の測定を行うことで,絶対年代や相対年代の見積もりやメタン湧出現象の確認が可能となります.上越沖では,約2万年前の最終氷期最盛期(Last Glacial Maximum;LGM)において,メタンハイドレートの大規模な分解が起き,メタンが大量に噴出した結果,底生有孔虫群集組成底生生物環境が局所的に変化した可能性が指摘されています.
2-2.近年の研究成果
Ohi et al.(2007)やUsami et al.(2013)の研究では,コア中に多く分布する薄いラミナを伴う暗色層において貧酸素に耐性を持つ種が多産することが認められました.特に,LGMにおける厚い暗色層の群集は底生有孔虫が全く産出せず非常に特徴的で,当時の日本海底層が底生生物が棲めないほどの無酸素環境であったことを表しています.一方,上越沖の海鷹海脚ではLGMにおいても底生有孔虫が僅かながら産出するという中川ほか(2009)の結果を確認でき(Mahsa et al., submitted),加えて,メタン湧出と関連が深い(秋元ほか,1996など)有孔虫種R. cornutaが増加する層準が見つかりました(未公表データ).
2-3.今後の展開
近年の研究より,LGMにおいて特殊な底層環境変化のイベントが発生したという観察結果が報告されています.しかしながら,LGMにおけるメタンハイドレート層の不安定化の要因やそれが地球環境に及ぼした影響もしくはその有無などについては,未だ明らかにされていません.また,この現象についての報告は上越沖にのみに限られており,まだその他のハイドレート分布域では検討されておりません.したがって,まず上越沖における地質学的な検討をより詳細に行い,その他の海域と比較検討していきたいと考えています.
地下微生物とガスハイドレート
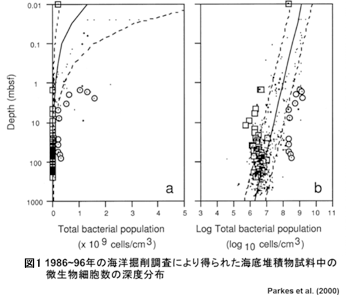
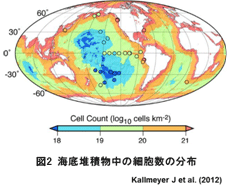
1986年のペルー沖で行われた海洋掘削調査 (ODP112) をはじめとして行われてきた海底掘削調査により、海底表層から深度500 mの範囲の海底堆積物中に1 cm3あたり数百万から数十億細胞 (106~109 cells/cm3) の微生物が生息していることが明らかになった (図1)。海は地球表面の70%を占めることからも、海底には無視できない量のバイオマスが存在していると言われている (図2)。
 大きなバイオマスが存在するような海底堆積物中では、太陽エネルギーによる光合成を利用して作られた有機物やその分解生成物に依存した生態系がひろがっている。その一方で、地球内部の無機化合物の化学反応によりエネルギーを獲得する化学合成生態系の存在も知られている。例えば、地下深部由来の水素と二酸化炭素からメタンを合成するメタン生成菌が一次生産を支えることがある。
大きなバイオマスが存在するような海底堆積物中では、太陽エネルギーによる光合成を利用して作られた有機物やその分解生成物に依存した生態系がひろがっている。その一方で、地球内部の無機化合物の化学反応によりエネルギーを獲得する化学合成生態系の存在も知られている。例えば、地下深部由来の水素と二酸化炭素からメタンを合成するメタン生成菌が一次生産を支えることがある。
近年のDNAを用いた分子生物学的手法の発達により、地下環境には系統学的にも生理学的にも既知の微生物とは異なる微生物が存在していることがわかってきている。また、地下環境では、生物が利用できるエネルギー源が限られており、エネルギー源の供給は低く、原位置での微生物代謝活性は低いと考えられている。大腸菌が20分で倍に増えるのに対し、地下微生物は数日から数年、遅いものは数百年以上かかって数が倍になると推測されている 。
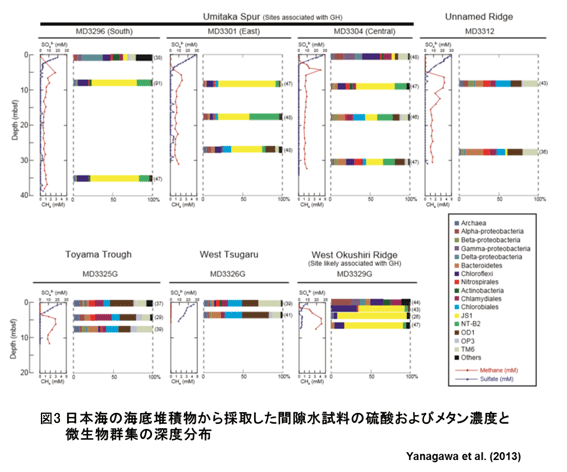
一般的に、海底堆積物中の硫酸塩濃度は、表層から海底深部に向けて減少し、深部では枯渇する (図3)。その硫酸塩が枯渇する深度より深部で、メタン生成古細菌の活動が活発に起こり、メタン濃度が上昇する。硫酸塩枯渇深度を境界 (Sulfate-Methane Interface, SMI) として、浅部でのメタンを消費する嫌気的メタン酸化細菌からメタンを生成する古細菌へと微生物活動がシフトする。SMI深度は、表層からの硫酸濃度の拡散と深部から浅部へのメタンの拡散、微生物によるメタンや硫酸の消費活動とメタン生成活動などに依存するため、サイトによって異なる (図3)。
海底堆積物中で微生物により生成されるメタンはメタンハイドレートの起源となりえること、メタンの生成と分解反応はいずれも古細菌の活動によって媒介されることが知られており、メタンハイドレートの集積や分解過程において微生物活動の寄与が考えられる。したがって、今後のメタンハイドレートの開発利用のために、メタン生成や分解に関わる微生物について生態と機能、微生物活動の定量的な評価が必要とされる。これまでもメタンハイドレートを胚胎する堆積物試料を対象に微生物群集の解析が行われてきたが、得られたDNA配列データを既存のデータベースと照合すると、海底で優占する微生物の多くが、系統学的に新しく、未培養の性状未知の微生物であり、それらの微生物の生態や機能についての評価は不十分である。Yanagawa et al. (2013) によると、日本海のメタンハイドレート胚胎試料では胚胎していない試料と比べ、未培養系統群JS1とNT-B2 (図3の黄、緑) が優占していた。JS1のグループは、海底堆積物で普遍的にみられる系統であるが、近縁な系統が冷湧水やペルー沖や南海トラフのメタンハイドレート胚胎試料での優占が明らかにされており、その分布はメタンガスの豊富な環境と関連するのではないかと推測されている。
ガスハイドレート研究所では、ハイドレート胚胎堆積物を用いて微生物学的解析を行い、メタン生成菌やメタン酸化細菌などのメタンの挙動に関与する微生物群集の分布を明らかにし、メタン酸化・分解活性の定量的な評価を行っている。微生物の生態や機能を明らかにすることは、微生物の物質循環への寄与を解明することにつながる。最終的には、ガスハイドレートの資源ポテンシャル評価のため、ガスハイドレートの起源や集積および分解メカニズムの解明、メタンの挙動に伴う環境影響評価を行うことを目標としている。